Biological Survey of Ironwood Forest National Monument
Natural History of the Desert Ironwood Tree (Olneya tesota)
Photographs by Mark Dimmitt

Flowering ironwood tree, Saguaro National Park

Ironwood in lush foliage, Ironwood Forest National Monument
Synopsis of published literature
Compiled by Tani Hubbard
The tree known in the U.S./Mexico borderlands as desert ironwood or palo fierro (Olneya tesota) is one of many woody legumes found in washes and hillside drainages in the Sonoran Desert. It ranks among the most ecologically and economically important plant species in the region. Ironwood functions as a “nurse plant” and a “habitat-modifying keystone species” of benefit to many other species of flora and fauna. While Ironwood is not endangered or threatened, its populations dwindle annually over tens of thousands of square kilometers.
Ironwood is nearly endemic to the Sonoran Desert (Turner et al. 1995). The species was first described in 1854 as the sole species of the genus Olneya by botanist Asa Gray and is still recognized as a monotypic genus (Lavin 1988). Ironwood is similar in morphology to only two other legume genera, peteria (Peteria sp.) and brushpea (Genistidium sp.). While all three genera have narrowly elliptical leaves and less than 12 ovules per pod, Olneya is distinguished by its paired leaves, flower clusters on short shoots that extend from the middle of the stem (instead of the end of a branch), and pods more rounded than the pods of peteria and brushpea.

Ancient ironwood stump
Ironwood as a species may have evolved as the Sonoran Desert flora formed in the middle Miocene (ca. 15 to 8 million years ago) (Van Devender 2000), but most paleogeological records of Ironwood date from the mid- to late Holocene. Dating of ironwood trees is difficult through standard tree-ring dating, but annual trunk diameter growth rates (Turner 1963; Suzán 1994) and unpublished radiocarbon dating estimates (Suzán 1994) suggest that some trees have persisted for more than 800 years. The wood of the Ironwood is one of the hardest and heaviest woods in the world (Búrquez 1999). It is remarkably resistant to rotting, perhaps because its heartwood is rich in toxic chemicals that make it essentially non-biodegradable (Dimmitt 2000a). Ironwood trunks can persist for up to 1600 years (Dimmitt 2000a). (See further discussion of ironwood longevity below.)
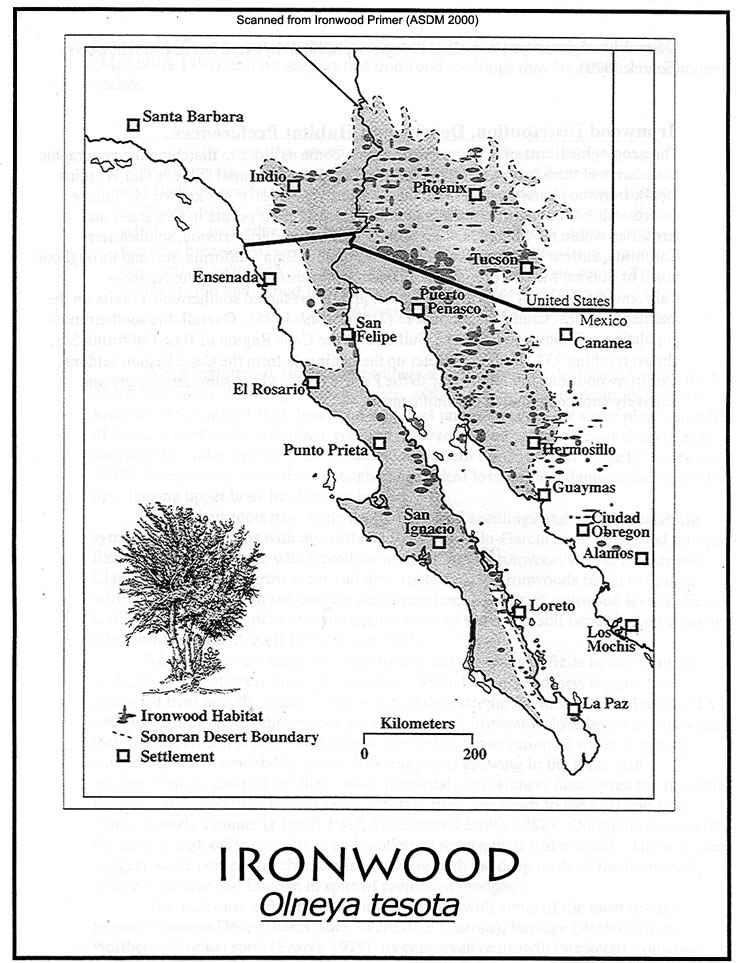
Distribution map of Ironwood Trees
Map courtesy Bill Singleton
Pima County (AZ) Administrative Office
Geography
The geographic limits of ironwood distribution are closely matched with the boundaries of the Sonoran Desert. Ironwood barely reaches into adjacent Mohave desertscrub, coastal thornscrub south of Guaymas, Sonora, and foothills thornscrub east of Hermosillo, Sonora. It occurs in five states and territories within the Sonoran Desert region: southwestern Arizona, southeastern California, eastern Baja California, Baja California Sur, and Sonora, Mexico. Populations occur from sea level to 1100 m (3280 ft) in elevation, where low winter temperatures and catastrophic freezes limit its distribution. Near its northern limit ironwood grows best on rocky benches and slopes, above the valley bottoms that characteristically have cold air pockets at night that would damage leaves and young branches (Turner et al. 1995). While ironwood occurs in all six subdivisions of the Sonoran Desert, it varies greatly in its density and relative dominance among these regions. In the U.S., the highest ironwood densities recorded per hectare are in Arizona Upland sites in Pima County (Ragged Top, 35 trees/ha = 14.2/acre; Cocoraque Butte [Roskruge Mountains] and Saguaro National Park West, 22 trees/ha = 8.9/acre) (ASDM 2000). In the IFNM survey some of our 0.4-hectare census plots had much higher densities that would extrapolate to more than 300 ironwoods per hectare. Ironwood densities are much lower in Mexico (mean density of 6.6 trees/ha = 2.4/acre across many plots in coastal and central Sonora). Elevational range is also greater in the species’ northernmost limits in the Arizona Uplands and Lower Colorado River Valley.

Dense Arizona Upland forest on granite
near Ragged Top, Ironwood Forest National Monument
The densest stands of ironwoods and palo verdes occur where the soil is derived from Precambrian Oracle granite. This granite is characterized by large crystal size and it weathers into a coarse, very porous soil that allows deep infiltration of water and air. This porous, well-aerated soil permits tree roots to penetrate deeply to reach the deep moisture. Soil explains much of the lushness of the tree growth in the Silver Bell region. The reason for the greater diversity of plants associated with ironwood trees here compared with other regions of the Sonoran Desert is not known.
Characteristics, Phenology, and Physiology
Ironwood may take the shape of either a multi-trunked shrub no more than two meters in height, or a canopy-forming tree with one thick trunk achieving heights up to 15 meters (49 ft; Shreve and Wiggins 1964, Solís-Garza 1993, Arizona Register of Big Trees 2000). The largest known ironwood, located close to Gila Bend, measures 4.32 meters (14.2 ft) around its trunk, with a canopy height of 15 meters 49 ft), and a crown spread of 14 meters (46 ft; Arizona Register of Big Trees 2000). The ironwood “leaf” is doubly divided into 4 to 12 pairs of narrowly elliptic leaflets called pinnae. Each leaf consists of two to four “fingers” with paired leaflets down the sides of each. This compound leaf has a pair of small curved spines at its attachment to the branch (Dimmitt 2000a). These leaflets have a bluish-green cast to them, producing a mottled canopy quite unlike the yellow-green of mesquites (Prosopis spp.) and palo verdes (Parkinsonia spp.) growing in the same region. Ironwood’s clusters of flowers bloom on the end of short shoots along the branches (Lavin 1988).

Ironwood trees flowering at the Arizona-Sonora Desert Museum (Tucson Mountains)

In Tucson, ironwood flowers and fruit occur in most years, but are abundant only four years per decade (Dimmitt 2000a). This variable level of flowering and fruiting, along with differences in rainfall each year, may cause a pattern of mass seed production and seedling germination that occurs as occasional bursts. Suzán (1994) observed this unpredictable pattern of germination, known as “discrete episodic recruitment.” Flowering and fruiting require considerable diversion of nutrients and energy from other parts of the plant. Branches that produce flowers often drop their leaves during bud formation, and re-leaf when summer rains begin. In some years flowering does not occur at all (Dimmitt (2000b). Patterns of flowering and fruiting generally occur in a south to north wave. Flowers and fruit occur as early as March in the southern states of Sonora and Baja California, Mexico, than in Arizona and California to the north.

Ironwood leaves usually turn yellow and are shed
in April before the trees flower.
The flowering period in each locality lasts only 10-18 days. These flowers attract one generalist bee and two solitary specialist bees. After pollination occurs, ironwoods produce slightly curved, knobby pods that reach lengths of 3-6 cm (1.2-2.4 inches) and widths of 8-9 mm (ca. 0.4 inch). These pods contain one to eight ovoid, shiny, coffee-colored, and extremely hard-shelled (at maturity) seeds (Solís-Garza and Espericueta 1997). Seed maturation coincides with the summer rains, increasing the probability of immediate germination (Shreve and Wiggins 1964). Maturation occurs within four to eight weeks of pollination (late June through August) (Turner et al. 1995). The seeds are small and light compared to those of blue palo verde or the indehiscent pods of mesquite (4,440-4,480 seeds per kg. Dry weight) (Kraugman 1948). They are high in protein and soluble fiber, but they also contain bitter chemicals that serve as deterrents to herbivores, reducing palatability and digestibility.

Ironwood tree in fruit.
Ironwood seeds mature at a time when little else is producing fruit in the Arizona Upland region (Dimmitt 2000b), leading to a high dependence of wildlife on the seeds. Many animals gather and store ironwood seeds in caches to be eaten later. Roughly half of all new germinated seedlings found for plants such as jojoba (Simmondsia chinensis) and palo verde occur in tight clusters near rodent burrows (McAuliffe 1990). While studies have not been conducted for ironwood, it is likely that ironwood seedlings germinate from rodent caches.
Ironwood grows extremely slowly, perhaps due to its low rates of photosynthesis that keep it from wasting soil moisture. These slow rates of biomass accumulation contribute to the remarkable density of its heartwood. Like other desert legume trees, ironwood trees conserve water during the high daytime temperatures and during dry seasons. Ironwoods lose less water through their leaves than other woody perennial plants. During long droughts, the trees slough off leaves, limbs and rootlets to reduce their water needs. The water use efficiency of ironwood ranks with some of the most drought tolerant Sonoran Desert plants, such as creosotebush (Larrea), bursage (Ambrosia), and wolfberry (Lycium spp.) (Szarek 1979). Considering ironwood’s conservative growth rate, small leaves, diffuse canopies, and preference for arid and hyper-arid xeroriparian soils, it is not surprising that they exhibit relatively low levels of annual net primary productivity (55 g. dry weight/m2/yr. = 15.6 oz/yd2/yr) as well as low gross productivity (7.42 kg = 16.3 lb. Dry weight/tree/yr) (Szarek 1979).
Ecological Importance

This ironwood tree founded a minicommunity. The young palo verde in the foreground and the large saguaro (as well as smaller plants not visible) established beneath its canopy. Now several small saguaros and shrubs are growing in the shelter of the palo verde. The adjacent ground is comparatively open. Tucson Mountains, AZ.
The ecological importance of the ironwood tree comes largely through the roles it plays for over 500 other species in the Sonoran Desert (Arizona-Sonora Desert Museum 2000). Ironwood trees function as a habitat-modifying keystone species, that is, a species that exhibits strong influences on the distribution and abundance of associated species (Mills et al. 1993). A chain of influences generated by ironwoods on associated understory plants affects their dispersal, germination, establishment, and rates of growth as well as reproduction. These ecological dynamics are termed “nurse plant ecology.” Other large trees co-occur with ironwoods along washes, but ironwoods may be the only tall branching woody plants on the valley floors or bajada slopes (Vander Wall 1980). Their relative influence on plant and wildlife diversity is proportionally greater in plains and rocky slope habitats above ephemeral and intermittent watercourses. Along watercourses, ironwood is but one of many nurse plants available. In addition, the size and foliar density of an ironwood are strong factors influencing their relative value as nurses. Medium-sized mature ironwoods harbor a greater diversity of understory plants than either ironwood saplings or the largest, ancient shade-forming ironwoods (Tewksbury and Petrovich 1994, Suzán et al. 1996). Some mid-sized trees, however, do not necessarily serve as nurses for many plants, especially if grazing is heavy.

Saguaro seedling beneath a palo verde tree.
Mesquite and palo verde also serve as nurse plants, however, each tree caters to slightly different sets of plants in its “nursery.” Ironwood is the dominant nurse plant in some subregions of the Sonoran Desert. As nurse plants, ironwoods provide safe sites for seed dispersal, protect seedlings from extreme cold and freezes, protect saplings from extreme heat and damaging radiation, and function as prey refugia. Also, like other legumes, they alter the soil composition beneath their canopies, enriching the soil with nutrients such as nitrogen.

The ironwood (right-center) is several times older than the same-size foothill palo verde to the left. The ironwood also has a denser canopy (next paragraph).
An ironwood canopy typically has been functioning as a safe site for seed dispersal for three to four times longer than mesquite or palo verde canopies of the same volume. The long life span of ironwood trees and the stability of the microenvironments they create increase the probability that seeds might be dispersed to these “safe sites” for germination and establishment (Tewksbury and Petrovich 1984). Due to the fact that ironwoods tend to be the tallest trees in desertscrub and xeroriparian vegetation (Vander Wall 1980), they function as the primary roosts in their landscape for both local breeding and migrating birds. Ironwoods and their nurseries make the structure of vegetation much more diverse providing birds with more nesting opportunities. Birds in turn generate a literal “rain” of seeds and whole fruit. Partially digested fruit from this “rain,” or from defecation of other animals, are torn apart by animals seeking to gain sustenance while selecting out toxic or distasteful portions of the fruit. Seeds also flow into the areas underneath ironwoods during storms and floods where they are trapped by exposed tree roots, or by the stems, roosts, and litter of understory herbs, vines and shrubs.
Ironwood canopies provide microenvironments buffered from freezes for understory plants. Suzán (1994) determined the winter microenvironments under mature ironwood trees may be 4 C (6.6° F) warmer than adjacent open environments and 1 C (2.8° F) warmer than under other vegetation. Studies of cactus seedling vulnerability demonstrate that without the protective cover of desert legumes, the distributional ranges of saguaro (Carnegiea gigantea), organ pipe (Stenocereus thurberi) and senita (Lophocereus schottii) would retreat many kilometers to more southerly, frost-free areas (Nobel 1980). In addition to frost protection, nurse plant canopies provide relief from heat and radiation stress. They reduce the exposure that leads to tissue damage and destruction of understory seedlings and saplings (Suzán 1994, Tewksbury et al. 1998). When stripped of ironwood’s protective cover above them, some cacti actually suffer sunburn and die (Nabhan and Suzán 1994). Where open soil temperatures can reach 65 C (148°; F), the 15 Centigrade degrees (27 Fahrenheit degrees) cooler temperatures under ironwood canopies increase seedling survival rates and decrease water stress in mature plants (Suzán 1994).

This exposed juvenile saguaro has been eaten by a jackrabbit (see scat pellets around base). If it were growing under a larger nurse plant than this shrub, it would probably have escaped predation.
In addition to serving as a buffer from abiotic stresses, ironwood buffers nursery plants from some, but not all, biotic stresses impacting their survival and reproduction. McAuliffe (1984a) demonstrated spiny, or thorny nurse plants can dramatically reduce predation on cactus seedlings by large and small herbivores, such as ungulates, rabbits, and rodents. The spiny, low-sweeping branches of the ironwood provide an effective prey refugium for vulnerable seedlings intertwined within its foliage. However, seedlings not fully sheltered by down-sweeping branches can suffer higher levels of predation due to the resting, nesting, or burrowing of desert tortoises, rabbits, jackrabbits, and packrats under ironwoods (McAuliffe 1984a).
Legumes such as ironwood and mesquite influence the soil composition beneath their canopies in several ways (Garcia-Moya and McKell 1970). Ironwoods “fix” nitrogen through symbiotic relationships with Rhizobia bacteria (Felker and Clark 1981). They also “pump” nitrogen and other nutrients up from their deepest root zones. Ironwoods incorporate these nutrients into their foliage, over time enriching topsoil composition as their fallen leaves gradually accumulate and decomposes beneath their canopies. Ironwoods and mesquites also act as traps for the nutrient-rich organic debris carried by flash floods (Nabhan 1993). The “resource islands” around ironwood and mesquite trunks support high densities of symbiotic bacteria and fungi that aid in the establishment of understory plants, providing them with moisture and nutrients not available in barren interspaces. The differences in the mycorrhizal fungi and soil composition under ironwoods and mesquites allow them to favor different sets of understory plants creating heterogeneity through “patch dynamics.” Ironwoods tend to slightly increase alkalinity and moisture availability, hardly effect soil texture, but significantly increase root, bacteria, and fungi densities where mesquites decrease soil alkalinity and increase clay content and moisture availability.
Threats

Ironwood Forest National Monument viewed from Saguaro National Park to the west. Agriculture and urbanization in the intervening Avra Valley could isolate the two, fragmenting the habitat into smaller parcels that will not support some wide-ranging species such as mountain lions. The Waterman Mountains are on the middle horizon, with the Silver Bell Mountains and Ragged Top to the right. The Roskruge Mountains are just visible on the far left horizon, and the taller Santa Rosa Mountains on the Tohono O'odham Nation to their right.
Ironwood habitat faces threats from habitat fragmentation due to urbanization and conversion of natural habitat to agricultural lands. The population explosion in the Sonoran Desert over the past 50 years has also led to increasing recreational impacts in ironwood habitat. There are also preliminary indications that both woodcutting and buffelgrass competition can decrease understory species richness and diversity (Suzán 1994, Búrquez and Quintana 1994). Ironwood cutting can result in greater damage to understory plants (Nabhan and Suzán 1994, Suzán et al. 1999). Nurslings exposed by woodcutting have a greater probability of damage from radiation, breakage from trampling, and death due to browsing (Nabhan and Suzán 1994). Solís-Garza and Espericueta (1997) have confirmed that virtually no ironwood regeneration had occurred to date in areas where commercial woodcutting has been permitted in Sonora. Buffelgrass is highly invasive, decreases plant species richness and diversity in native plant communities, and increases fire frequency. Fires in communities invaded by buffelgrass tend to be hot burning and destroy ironwood and other trees, shrubs, and cacti.
While ironwood is not considered endangered because of its large range, it is easily overexploited because of certain life history traits, primarily its slow growth rates and low levels of seedling establishment (Suzán 1994). Ironwood populations play a vital role in sustaining other species and populations of the Sonoran Desert. If ironwoods were eliminated from Sonoran Desert habitats, there would be a decrease in the density of associated plants and subsequently in associated local faunal communities. Ironwoods must be protected both to maintain the diversity and lushness of the Sonoran Desert communities they inhabit and to maintain the regeneration dynamics of rare plant populations that grow under its canopies. Ironwoods are truly a hallmark of the desert landscape living well beyond other desert plant species. The ironwood is both a constant witness to a changing environment and an active participant in the maintenance of generations of lush Sonoran Desert plant and animal communities.
Ecology of Ironwood Trees in Ironwood Forest National Monument
by Mark Dimmitt

All of the trees in the background are ironwoods. Most of them are growing in small drainages on the finely-dissected bajada and valley floor. Little Maria Mountains northwest of Blythe, California.

This large wash in the Chuckwalla Valley west of Blythe, California is vegetated with ironwood trees (the darker ones), some blue palo verdes (Parkinsonia florida), and a few desert willows (Chilopsis linearis) and smoke trees (Psorothamnus spinosus). The McCoy Mountains are on the horizon.
In most of the Sonoran Desert ironwood trees grow mainly on valley floors and are restricted to washes in the driest habitats (Turner et al. 1995; images above). Arizona Upland is the highest elevation, wettest, and coldest of the six subdivisions of the Sonoran Desert (Shreve 1964). Ironwoods behave differently in this zone; they live on bajadas above the cold valley floors (image below left). On the eastern side of IFNM which is mostly Arizona Upland, ironwoods are abundant on most of the bajadas of the Roskruge, Silver Bell, Waterman, and Ragged Top ranges. Occasionally they are even common on rocky slopes such as on Cocoraque Butte in the Roskruge Mountains. Their eastern range limit is in the Tucson Mountains and the extreme southwestern foothills of the Santa Catalina Mountains. They extend into Avra Valley in some drainages, but almost not at all into the colder Santa Cruz Valley in the vicinity of Tucson.

Ironwood trees grow on the upper bajadas and lower slopes of the Tucson Mountains (above) and the eastern ranges of Ironwood Forest National Monument. This is atypical habitat for the species.

Ironwoods typically grow on valley floors, as here in the Avra Valley east of the Samaniego Hills in IFNM. There are few ironwood groves in Avra Valley, however, because of its higher elevation and more frequent frosts that kill this tree of tropical origin.
From east to west Ironwood Forest National Monument trends to lower, warmer, more arid terrain. The transition from AZU to LCV is apparent between the Silver Bell and West Silver Bell Mountains (maps below). It is quite obvious by the time one reaches the Sawtooth Mountains in the far northwest part of the National Monument; there the slopes are thinly vegetated with trees and saguaros and trees on the valley floors are mostly restricted to washes. Along this transect ironwood trees change their habitat from their anomalous location on upper bajadas in the east and move down into their more characteristic habitat of valley floor washes in the west. However, they are not ubiquitous in washes. Ironwood trees are abundant in some washes, while adjacent ones only half a mile away are dominated by mesquite or blue palo verde and nearly or completely devoid of ironwoods.
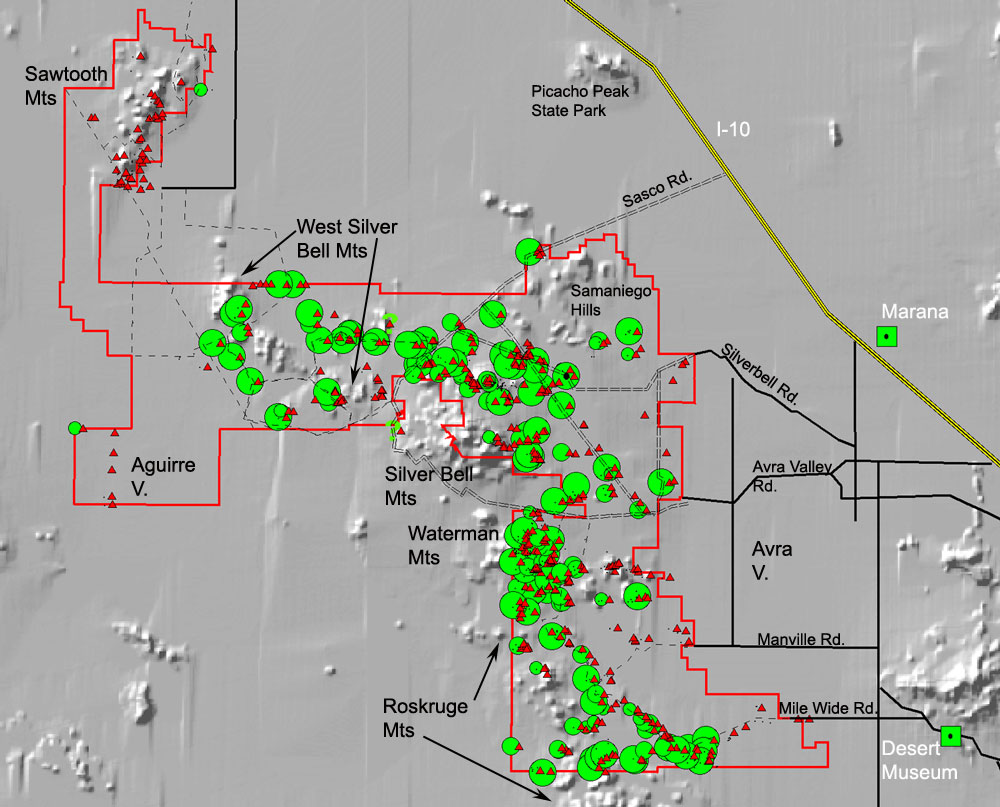
Distribution of desert ironwood trees within Ironwood Forest National Monument. The size of the green circles indicates the relative abundance of ironwood trees from rare to common. The red triangles are sites surveyed. In the east ironwood trees are common on lower slopes and bajadas and not on valley floors. In the West Silverbell Mountains they are absent from most slopes but common on the lower bajadas, and the Sawtooth Mountains area is almost devoid of ironwoods in any habitat.
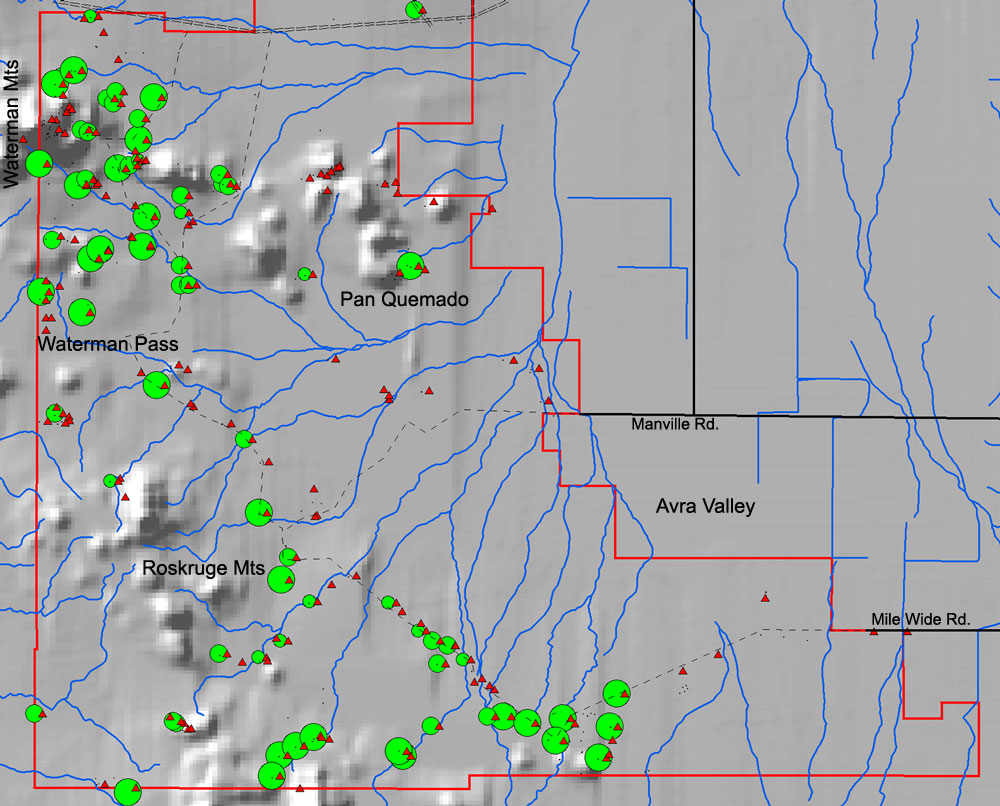
In the eastern part of IFNM ironwood trees occur on bajadas and lower mountain slopes in Arizona Upland and are not restricted to washes (blue lines). They are excluded from most of Avra Valley by winter cold.

Ironwood tree distribution in the northeastern part of IFNM is similar to that in the southern part. There are a few groves of large trees in parts of Avra Valley, especially east of the Samaniego Hills.

In the midwestern part of IFNM ironwood trees more often occur in their typical habitat - large washes on valley floors and lower bajadas. Valley floors are lower elevation and thus warmer. They are nearly absent from the slopes and bajadas of the arid West Silver Bell Mountains. Aridity would be the logical reason, except that less drought-tolerant foothill palo verdes are common on these slopes. There are good groves on some bajadas such as the south side of granitic Solo Peak (the rest of the West Silver Bells are other volcanics).

Large washes on low valley floors are the typical habitat of ironwood trees. Tiro Wash, West Silver Bell Mountains, Ironwood Forest National Mon.
Ironwood tree distribution extends all the way to the western margin of the Sonoran Desert near Palm Springs, California. But we found no ironwood trees in the Sawtooth Mountains and adjacent valley floors within the IFNM boundary. (Two trees grow at the edge of a quarry just outside the boundary.) Apparently the drainages in this region do not collect sufficient runoff to support them.
Ironwood longevity
On most of the rocky bajadas ironwoods are small trees or large shrubs seldom more than four meters (13 ft) tall and much shorter than the saguaros that usually grow with them. A large proportion of the individuals in these habitats have old, dead trunks that have resprouted from the crowns. Some have evidently died to the ground (topkilled) at least twice in the past. So even though most of the trees on these rocky sites are rather short and look like saplings at first sight, many are in fact ancient (see images below).

This ironwood in the Roskruge Mountains (Ironwood Forest National Monument) appears to have died to the ground and crown-sprouted at least twice.

This ironwood tree in the Chuckwalla Valley, California shows evidence of four generations of topkill and resprouting.

Ironwood trees that grow in deep soil in large washes are probably not very old. This one is east of Indio, California.
Comparison of ironwoods with foothill palo verdes in the same location provides further evidence of the longevity of ironwood trees. A significant percentage of foothill palo verdes died during the droughts of the mid 1990s and 2001, while we found almost no ironwood trees that had died or were topkilled recently. The droughts that caused the observed topkill in the ironwoods must have been more severe than what we have recorded during the last century, and the same drought would likely have killed most of the mature foothill palo verdes (Bowers and Turner 2001). Because mature foothill palo verdes in the nearby Tucson Mountains are 125 to 175 years old (Turner et al., 2003), the mature palo verdes currently living in the Monument are presumably at least 100 years old. Therefore the younger ironwood stumps visible at present must have been topkilled before today’s mature palo verdes established, and those that have topkilled and resprouted twice or more are probably several centuries old. Conversely, the large, vigorous specimens in well-watered washes may be only 100-200 years old.
 The heartwood of Olneya tesota is almost nonbiodegradable. The minerals deposited in the heartwood that make it too dense to float in water are toxic to most saprophytes and termites. This ironwood tree stump is weathering away by physical processes. It probably died two or more centuries ago. Chuckwalla Mountains, California. |
The most likely candidate for the last catastrophic drought that killed ironwood trees to the ground is the one of 1891-1904; some call it the most severe drought documented in southern Arizona (Turner et al. 2003). But others consider the mid century drought of 1942 to 1977 to be the worst since 1700 A.D.; it eliminated most pinyons and junipers established before 1850 in much of New Mexico (Swetnam and Betancourt 1998). That drought also thinned out many desert landscapes (Turner et al. 2003). A more recent drought in the 1990s killed significant numbers of palo verdes (Bowers and Turner 2001). We can still identify the carcasses of these palo verdes at the Arizona-Sonora Desert Museum in the Tucson Mountains. We observed no mortality of ironwood trees in undisturbed areas of the Tucson Mountains during that period (Dimmitt pers. obs.), nor does the literature reviewed mention dieback of ironwood trees during the past several decades.
The primary cause of death of ironwood trees is unknown. If drought is a significant mortality factor, then these events must be spaced centuries apart. There is a root rot caused by a fungus in the genus Ganoderma (Olsen 1999, Hine 1999). We have observed its effects of slow yellowing and diminution of foliage over several years at ASDM and at Bach’s Greenhouse Cactus Nursery in the foothills of the Tortolita Mountains. We did not observe these symptoms in any trees in IFNM, nor did we find any publications documenting its occurrence in wild trees. The disease seems to be associated with disturbance.
 |
 |
| Considering how long dead ironwood trees persist in the landscape, mortality must be extremely low because it is rare to see more than a few dead trees in a population. A disturbing exception is these old ironwood trees in the Chuckwalla Valley (west of Blythe, CA) that have been declining for at least 30 years. In some areas more than three-quarters of them are now dead. This mortality has not been reported in the scientific literature. It may be long-term drought, though groundwater pumping has also been suggested. Another possibility is that the smaller drainages were diverted into larger ones that had culverts under the old highway (now replaced by Interstate 10 a few miles to the south and renamed Ford Dry Lake Rd. and Corn Spring Rd.). The summer storm (right) came much too late to save the tree in the foreground, though the large wash in the background has healthy trees that have leafed out from an earlier storm. | |
 The largest ironwood known in Ironwood Forest National Monument. |
The largest ironwood tree we found in the Monument is 11 meters tall and 13 m wide (36 X 43 feet) in Avra Valley east of the Samaniego Hills. This area is characterized by sparse but large ironwood trees; the plot had ten trees over 7 m (23 ft) tall.
It is worth noting that Ironwood Forest National Monument was not created and named after the ironwood tree because it has the largest trees or densest forests. The Monument’s claim to fame is that in this area ironwood trees have more ecological associates than anywhere else this phenomenon was measured (ASDM 2000).
 Three generations of a keystone species. The old ironwood stump that died several centuries ago may be a grandparent of the 200-year-old tree in the background, which may in turn be a parent of the two-week-old seedling in the foreground. Bajada of Cottonwood Canyon, south of Joshua Tree National Park, California. |
http://desertmuseumdigitallibrary.org/programs/ifnm_ironwoodtree.php